شنبه, ۱۵ اردیبهشت, ۱۴۰۳ / 4 May, 2024
مجله ویستا
نگاهی به مهندسی تولید متابولیت های ثانویه
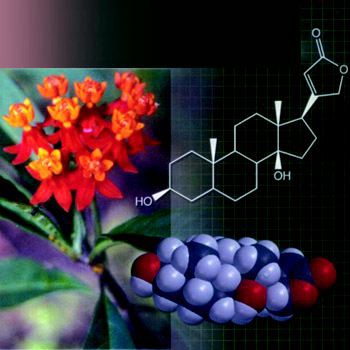
• تفاوت های متابولیت های ثانویه با متابولیت های اولیه
سلول ها خصوصاً سلول های گیاهی دو دسته از تركیبات را تولید می كنند؛ متابولیت های اولیه و متابولیت های ثانویه.
• متابولیت های اولیه
متابولیت های اولیه مستقیماً در رشد و متابولیسم درگیر هستند و شامل كربوهیدرات ها، لیپیدها، پروتئین ها و اسیدهای نوكلئیك می شوند. در گیاهان متابولیت های اولیه طی فرایند فتوسنتز تولید شده و سپس در ساخت تركیبات سلول نقش آفرینی می كنند. این تركیبات در حجم زیاد و با ارزش اقتصادی پائین تولید می شوند و عمدتاً به عنوان ماده خام صنعت، موادغذایی و افزودنی ها كاربرد دارند. روغن های گیاهی، اسیدهای چرب (برای ساخت صابون و شوینده ها) و كربوهیدرات هایی مانند ساكاروز، نشاسته، پكتین و سلولز مثال هایی از متابولیت های اولیه هستند. قیمت این قبیل تركیبات به طور میانگین دو دلار در هر كیلو بوده و تولید آنها در حجم انبوه امكان پذیر است. البته برخی از متابولیت های اولیه مانند میواینوزیتول و بتاكاروتن گران هستند كه علت قیمت بالای آنها، سختی استخراج و تخلیص آنها است.
• متابولیت های ثانویه
متابولیت های ثانویه از بیوسنتز متابولیت های اولیه به دست می آیند و به عنوان تركیبات فرعی و انتهایی متابولیسم اولیه در نظر گرفته می شوند. همچنین این تركیبات در فرآیندهای متابولیسمی وارد نمی شوند. مهمترین متابولیت های ثانویه آلكالوئیدها، فنولیك ها، روغن های ضروری، استروئیدها، لیگنین ها، تانن ها و فلاوونوئیدها هستند. متابولیت های ثانویه عمدتاً در گونه ها و خانواده های خاصی از سلسله گیاهان تولید می شوند. این تركیبات به مقدار كمی در سلول ذخیره شده و عمدتاً در سلول های تخصصی و در مرحله خاصی از چرخه زندگی گیاه تولید می شوند و همین امر استخراج و تخلیص آنها را در مقایسه با متابولیت های اولیه كه در تمام سلول ها تولید می شوند، دشوار می كند. گیاهان دارویی از لحاظ میزان متابولیت های ثانویه بسیار غنی هستند و تركیبات آنها را در انگلیسی Medicinal یا Officinal می نامند. این تركیبات كه از گروه متابولیت های ثانویه هستند، اثرات فیزیولوژیكی عمیقی بر پستانداران دارند و از مهمترین تركیبات دارویی هستند. از این گیاهان و اثرات فیزیولوژیك تركیبات موثره آنها به عنوان داروی خوراكی استفاده شده و در نتیجه این تركیبات، داروهای گیاهی یا داروهای طبیعی نام گرفته اند. استفاده از داروهای با منشاء گیاهی بدون انجام فرآوری خاصی، یعنی استفاده از پودر گیاه یا مواد موثره آن بدون خالص سازی عصاره گیاهی، از قدیم رواج داشته و حتی در حال حاضر و در پزشكی مدرن از طیف وسیعی از داروهای با منشاء گیاهی استفاده می شود. گرچه تعدادی از این داروها به طور مصنوعی برای مصارف ساخته می شوند ولی هنوز بسیاری از آنها از منابع طبیعی به دست می آیند. به محض اینكه اثر فیزیولوژیك یك گیاه دارویی خاص كشف شود، تلاش ها برای یافتن خصوصیات دقیق شیمیایی ماده موثره آن (داروی گیاهی) و در پی آن، یافتن روش تولید شیمیایی این تركیبات به طور تجارتی صورت می گیرد. برای تعیین خصوصیات شیمیایی و شناسایی یك متابولیت ثانویه، جداسازی آن به صورت كاملاً خالص الزامی و اولین قدم است. روش های جداسازی گوناگون و مراحل آن اكثراً طولانی است. متابولیت های ثانویه به صورت خالص و با نسبت های مشخص در پزشكی استفاده می شوند. البته در كنار خالص سازی مواد موثره گیاهان دارویی، آنها بدون تغییر نیز در سامانه های مختلف تهیه دارو استفاده می شوند. این نكته را نباید فراموش كرد كه در كنار متابولیت های ثانویه، تعدادی از متابولیت های اولیه نیز اثرات فیزیولوژیكی قوی دارند. اكثر این تركیبات پروتئینی بوده و عملكردهای مختلفی دارند. هورمون ها و زهرمارها مثال هایی از پروتئین های با اثرات فیزیولوژیكی قوی هستند. آنتی بیوتیك ها، واكسن ها و تعدادی از پلی ساكاریدها كه نقش هورمونی دارند، از جمله متابولیت های اولیه با اثر فیزیولوژیكی قابل ملاحظه هستند.
• مسیرهای تولید متابولیت های ثانویه
اسكلت كربنی متابولیت های ثانویه از كربوهیدرات ها تامین شده و طی فرآیند فتوسنتز ایجاد می شود. متابولیت های اولیه دیگری كه در تولید متابولیت های ثانویه نقش دارند، اسیدهای آمینه هستند. استیل كوآنزیم و اسید مِوالونیك نیز نقش مهمی در تولید انواع ترپنوئیدها دارند. مسیر اسید شیكیمیك نیز در تولید لیگنین ها و ایندول آلكالوئیدها نقش آفرینی می كند.
• مهندسی تولید متابولیت های ثانویه در گیاهان
در حال حاضر دستاوردهای زیاد و گوناگونی در مهندسی متابولیك متابولیت های ثانویه به دست آمده است. مسیرهای بیوشیمیایی مختلفی با استفاده از ژن های رمزكننده آنزیم های مهندسی شده و پروتئین های تنظیمی بررسی شده اند. در عین حال از ژن های آنتی سنس نیز برای مسدود كردن مسیرهای بیوشیمیایی و افزایش تولید متابولیت ثانویه خاصی استفاده می شود. متابولیت های ثانویه وظایف مختلفی در طول چرخه زندگی گیاه دارند. از جمله این وظایف انجام نقش تركیبات واسطه در برهم كنش گیاه و محیط اطرافش و برهم كنش گیاه با حشرات، ریزسازواره ها و حتی گیاهان اطراف است. تولید متابولیت های ثانویه همچنین ممكن است بخشی از سامانه دفاعی گیاه باشد. این تركیبات در تولیدمثل گیاه نیز نقش دارند كه از طریق جذب حشرات گرده افشان این كار را انجام می دهند. متابولیت های ثانویه گیاهی نقش مهمی در ایجاد كیفیت مواد غذایی (رنگ، طعم و بو) مختلف دارند. همچنین رنگ گیاهان زینتی و گلبرگ گل ها كه مهمترین خاصیت این دسته از گیاهان است، توسط متابولیت های ثانویه ایجاد می شود. بسیاری از متابولیت های ثانویه نیز برای تولید دارو، رنگ، حشره كش ها، طعم دهنده های غذایی و عطر استفاده می شوند. به دلیل كاربردهای فراوان، متابولیت های ثانویه موضوع جالبی برای تحقیقات اصلاح نباتات از طریق فنون مولكولی و مهندسی ژنتیك محسوب می شوند. در ده سال گذشته تحقیقات چندانی در ارتباط با متابولیت های ثانویه انجام نشده است. مانع بزرگ در انجام این تحقیقات اطلاعات اندك از مسیرهای تولید زیستی متابولیت های ثانویه و برهم كنش آنزیم های درگیر در این مسیر است. همچنین تعداد معدودی از ژن های مربوط به متابولیت های ثانویه در دسترس است. یكی از مسیرهایی كه مطالعات بیشتری در سطح ژن های دست اندركار آن نسبت به دیگر متابولیت های ثانویه انجام شده است، مسیر تولید فلاوونوئیدها و آنتوسیانین ها است. اكثر ژن های درگیر در مسیر تولید آنتوسیانین ها همسانه سازی شده و مطالعات فراوانی در سطح بیوشیمیایی، مولكولی و ژنتیك این دست از متابولیت های ثانویه صورت گرفته است. یكی از مهمترین دلایل مطالعات بیشتر در این زمینه، آسانی بررسی این مواد از روی رنگ گل ها به فنوتیپ قابل غربال گری است. دیگر مسیر تولید متابولیت های ثانویه كه در سطح آنزیم های درگیر و مواد واسط مورد مطالعه قرار گرفته است و اهمیت زیادی در داروسازی دارند، ایندول آلكالوئیدها هستند. اما در این مسیر هم ژن های محدودی مشخص شده اند. هدف از مهندسی ژنتیك مسیر یك متابولیت ثانویه، افزایش مقدار یك ماده خاص یا گروهی از تركیبات و یا حتی كاهش مقدار این تركیبات است. برای دستیابی به هدف دوم كه كاهش میزان تولید یك ماده خاص یا گروهی از مواد ناخواسته است، راه های مختلفی وجود دارد. [این مواد ممكن است تركیبات سمی در یك محصول گیاهی، مواد مانع خالص سازی یك فرآورده صنعتی یا موادی از این دست باشد.]یكی از این راه ها، مسدود كردن یك مرحله از مسیر تولید متابولیت ثانویه و مختل كردن تولید یا فعالیت آنزیم مربوط به آن مرحله است. این هدف می تواند با كاهش میزان mRNA مسئول تولید این آنزیم، با استفاده از فناوری آنتی سنس، RNAi یا بیان بالای یك آنتی بادی علیه آنزیم مسئول محقق شود. فناوری آنتی سنس به خوبی برای تغییر رنگ گل ها استفاده شده است. راه های دیگر دستیابی به این هدف، تغییر مسیر به سوی مسیرهای موازی یا افزایش كاتابولیسم ماده نهایی است. اما ممكن است هدف از انجام تحقیقات، افزایش تولید یك تركیب خاص در گیاه بوده یا انتقال ژن های مربوط به مسیر تولید یك متابولیت ثانویه به یك گیاه یا یك ریزسازواره مورد نظر باشد. همچنین ممكن است تولید یك ماده جدید كه به صورت طبیعی در گیاهان تولید نشود هدف یك پروژه تولیدی- پژوهشی باشد. در برخی روش ها با تغییر میزان بیان یك یا چند ژن، بر موانع تولید یك ماده غلبه می كنند و در روش های دیگر، با حذف مسیرهای موازی (رقابتی) یا كاهش كاتابولیسم ماده مورد نظر، مقدار آن ماده را در گیاه می افزایند. ایجاد تغییراتی در بیان ژن های تنظیمی كه كنترل مسیر تولید زیستی متابولیت های ثانویه را برعهده دارند نیز از جمله روش های افزایش یا كاهش تولید تركیب مورد نظر است. در ادامه این مطلب برخی از نتایج به دست آمده از بررسی نحوه تولید متابولیت های ثانویه ژن های مسئول و تنظیم بیان ژن های درگیر ارائه می شود.
• فلاوونوئید ها و آنتوسیانین ها
ـ ژن های مسئول بیوسنتز فلاوونوئید ها و آنتوسیانین ها
بیوسنتز فلاوونوئید و آنتوسیانین از اولین مسیر های متابولیت های ثانویه بود كه مهندسی ژنتیك آنها صورت گرفت. دلیل این موضوع این بود كه مسیر بیوسنتز آنها به خوبی شناخته شده بود و نتایج تغییرات انجام شده به راحتی از روی تغییر رنگ گل ها قابل مشاهده است.آزمایش های زیادی شامل افزایش بیان ژن های مختلف برای ایجاد رنگ های جدید گل یا تولید تركیبات جدید در گیاه انجام شده است.به خاطر خواص آنتی اكسیدانی این تركیبات افزایش میزان تولید و مقدار آنها در غذا های گیاهی موضوع جالب توجهی برای دانشمندان بوده است. اكثر پژوهش های انجام شده در زمینه بیوسنتز فلاوونوئید ها و آنتوسیانین ها در گیاه گوجه فرنگی صورت گرفته است. مشخص شده است كه آنزیمی از گروه آیزومراز به نام CHI از آنزیم های مراحل اولیه مسیر تولید فلاوونوئید نقشی كلیدی در افزایش تولید فلاوونول دارد. افزایش بیان ژن CHI گیاه اطلسی باعث افزایش سطح فلاوونوئید در پوست گوجه فرنگی به میزان ۷۸ برابر می شود همچنین با تولید گیاهان گوجه فرنگی اصلاح ژنتیكی شده، میزان فلاوونوئید آنها بیش از ۲۰ برابر گیاهان شاهد بوده است. این موضوع حاكی از آن است كه افزایش تولید تركیبات مفید برای سلامتی در محصولات حاصل از گوجه فرنگی امكان پذیر است.بیوسنتز ایزوفلاوون ها كه تركیباتی ضدمیكروبی هستند، بر اثر آلودگی میكروبی در حبوبات القا می شود. افزایش بیان ژن ایزوفلاوون سنتتاز كه یك آنزیم سیتوكروم P۴۵۰ است، باعث تولید این تركیبات در آرابیداپسیس، توتون و ذرت كه به طور عادی توانایی تولید این مواد را ندارند، می شود. تولید این تركیبات در این گیاهان بستگی به در دسترس بودن پیش ماده های مسیر فنیل پروپانوئید دارد. القا یا مهندسی این مسیر می تواند در اصلاح و بهبود سامانه بیوسنتز ایزوفلاوون در گیاهان میزبان مهم باشد.
ـ ژن های تنظیم كننده، تولید فلاوونوئید ها و آنتوسیانین ها
یك روش برای مهندسی بیان ژن های یك مسیر بیوشیمیایی استفاده از عوامل رونویسی خاصی است كه ژن های مسیر های بیوشیمیایی متعددی را كنترل می كنند. در دانه ذرت تولید آنتوسیانین توسط تركیبی از دو عامل رونویسی R و Cl تنظیم می شود. پروتئین های R شباهت زیادی با پروتئین هایی كه توسط ژن های مهره داران رمز می شود (ژن c-MYC) دارد در حالی كه پروتئین های Cl مشابه محصولات ژن های c-MYB است. القای مسیر تولید فلاوونوئید با افزایش بیان این دو عامل رونویسی R و C۱ در سلول های تمایز نیافته ذرت در محیط كشت صورت می گیرد. در برنج افزایش بیان این دو عامل رونویسی مربوط به گیاه ذرت در كنار یك ژن دیگر باعث فعال شدن مسیر بیوسنتز آنتوسیانین و همچنین افزایش مقاومت گیاه برنج به قارچ ها می شود. در آرابیداپسیس، یك عامل رونویسی از نوع MYB به نام PAP۱ تشخیص داده شده است كه افزایش بیان آن باعث تولید رنگدانه های بنفش پررنگ در زمان نمو گیاه می شود. این مثال ها نشان می دهد كه می توان با افزایش بیان یك یا چند عامل رونویسی (حتی با استفاده از ژن های بیگانه در گیاه میزبان)، بر این سامانه قدرتمند تنظیمی ژنتیكی در هنگام نمو گیاه، غلبه كرد. عوامل رونویسی گاهی به عنوان مانع ذخیره سازی تركیبات طبیعی نیز عمل می كند. غیرفعال سازی ژن عامل رونویسی MYB۴ در آرابیداپسیس می تواند باعث افزایش سطح موادی شود كه تحمل گیاه را نسبت به تشعشعات اشعه ماوراءبنفش می افزایند. افزایش بیان ژن های تولیدكننده پروتئین FaMYB۱ كه از دسته پروتئین های MYB است و از گیاه توت فرنگی به توتون منتقل شده است، باعث كاهش تولید رنگدانه های گل شده و سطح آنتوسیانین و فلاوونول در گیاه توتون پایین می آید. تحلیل این اتفاق اینگونه است كه FaMYB۱ در میوه توت فرنگی به عنوان یك مانع در برخی از مراحل مسیر تولید فلاوونوئید عمل می كند.
• آلكالوئیدها
ـ ژن های بیوسنتز ایندول آلكالوئیدها
مسیر ترپنوئید ایندول آلكالوئید موضوع بسیاری از تحقیقات مهندسی ژنتیك بوده است چرا كه حدود ۱۵ نوع از تركیبات این مسیر از نظر صنعتی اهمیت دارند كه آلكالوئید ضدتومور وینبلاستین از آن جمله است.
همه این آلكالوئیدها در یك قسمت از مسیر بیوشیمیایی تا تولید ماده واسط استریكتوزیدین مشترك هستند و از این نقطه، مسیر در گونه های مختلف گیاهی تولیدكننده مواد آلكالوئیدی منشعب می شود.بیشتر مطالعات انجام شده برای تهیه نقشه ژنتیكی بخش اولیه و مشترك مسیر و افزودن میزان بیان ژن های این بخش از مسیر بوده است. هدف از این كارها افزایش فعالیت مسیر تولید آلكالوئیدها است. به طور مثال، ژن های رمزكننده تریپتوفان دِكربوكسیدلاز (TDC) و STR-SS در سلول های گیاه Catharnthus roseus به خوبی مورد مطالعه قرار گرفته است. افزایش بیان TDC باعث افزایش ماده حدواسط تریپتامین شده در حالی كه به افزایش سطح آلكالوئیدها نمی انجامد اما با حضور STR، افزایش میزان آلكالوئیدها قابل ملاحظه است. با افزودن ماده حدواسط تریپتوفان و ترپنوئیدها به محیط غذایی لاین های سلولی مورد آزمایش، آنها ظرفیت تولید میزان بالایی از آلكالوئیدها را تا حدود یك میلی مول در لیتر بروز دادند. این موضوع به این معنی است كه شاخه ترپنوئیدی مسیر بیوسنتز متوقف شده است. مطالعات تكمیلی نشان می دهد كه مراحل محدودكننده دیگری نیز وجود دارند كه در اینجا از ذكر آنها خودداری می كنیم. TDC و STR در گیاهان دیگری كه مواد آلكالوئیدی تولید نمی كنند نیز بیان می شود. تغذیه كشت سلولی سلول های گیاه توتون با سِكولوگانین، باعث تولید استریكتوزیدین شده اما این ماده كه یك گلوكو آلكالوئید است، به جای ذخیره در واكوئل برای تولید ایندول آلكالوئید در مراحل بعدی (همانند آنچه در Catharnthus roseus اتفاق می افتد)، به محیط كشت ترشح می شود. این موضوع اهمیت جنبه های فیزیولوژیكی را در تولید زیستی متابولیت های ثانویه نشان می دهد به طوری كه نه تنها ژن های رمزكننده آنزیم های درگیر كاتالیز مراحل تولید زیستی در این امر مشاركت دارند بلكه ژن های دیگری نیز از جمله ژن های تنظیم pH و ژن های مربوط به انتقال مواد نیز یك پای قضیه هستند.
ـ ژن های تنظیم كننده تولید ایندول آلكالوئید
دو عامل رونویسی به نام های ORCA۲ و ORCA۳ كه در چندین مرحله از مسیر بیوشیمیایی تولید آلكالوئید در Catharnthus roseus نقش تنظیمی دارند، تعیین شده اند.افزایش بیان ORCA۳ به تنهایی باعث افزایش تولید آلكالوئیدها نمی شود چرا كه این عامل رونویسی، ژن G۱۰H را كه یك آنزیم سیتوكروم P۴۵۰ موثر در كاتالیز مرحله اول تولید سكولوگانین است، تنظیم نمی كند. اما پس از تغذیه سلول ها با پیش ماده سكولوگانین كه همان لوگانین است، تولید آلكالوئیدها تا میزان ۳ برابر افزایش می یابد. هر دو این عوامل تنظیمی (ORCA۲ و ORCA۳) درگیر تولید زیستی ترپنوئید ایندول آلكالوئید بوده گرچه كنترلی بر ژن G۱۰H ندارند. این موضوع نشان می دهد كه ژن های دیگری نیز درگیر تنظیم فعالیت این مسیر هستند كه می تواند موضوع پژوهش های بعدی در این زمینه باشد.نتایج تمامی این مطالعات یك پیام مهم را نیز داشت، برای افزایش بیان یك دسته از آنزیم های درگیر در یك مسیر بیوشیمیایی می توان از ژن های تنظیمی استفاده كرد به جای آنكه بیان هر یك از ژن ها را به طور جداگانه افزایش داد.
كسری اصفهانی
منبع : روزنامه شرق









































































































































































نمایندگی زیمنس ایران فروش PLC S71200/300/400/1500 | درایو …
دریافت خدمات پرستاری در منزل
pameranian.com
پیچ و مهره پارس سهند
تعمیر جک پارکینگ
خرید بلیط هواپیما
حماس دولت سیستان و بلوچستان انتخابات مجلس شورای اسلامی عراق چین دولت سیزدهم حسن روحانی روز معلم رهبر انقلاب معلمان
سیل هواشناسی ایران تهران آتش سوزی باران هلال احمر اصفهان سازمان هواشناسی شهرداری تهران معلم پلیس
قیمت خودرو خودرو ترکیه تورم مسکن قیمت طلا قیمت دلار بازار خودرو سهام عدالت دلار حقوق بازنشستگان ایران خودرو
مهران غفوریان تلویزیون ساواک سریال صداوسیما موسیقی مسعود اسکویی رضا عطاران تئاتر
موبایل
اسرائیل رژیم صهیونیستی غزه فلسطین جنگ غزه آمریکا روسیه اوکراین انگلیس نوار غزه ایالات متحده آمریکا یمن
فوتبال پرسپولیس استقلال لیگ برتر سپاهان باشگاه پرسپولیس علی خطیر باشگاه استقلال بازی تراکتور لیگ قهرمانان اروپا رئال مادرید
اپل هوش مصنوعی ناسا گوگل صاعقه تماس تصویری تلفن همراه مدیران خودرو
کبد چرب فشار خون طول عمر