شنبه, ۱ اردیبهشت, ۱۴۰۳ / 20 April, 2024
مجله ویستا
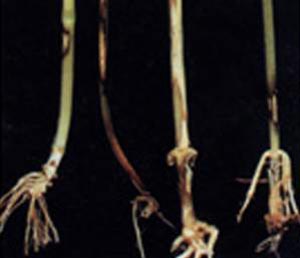
چگونگی مقابلهٔ گیاهان با تنش خشکی در مزرعه

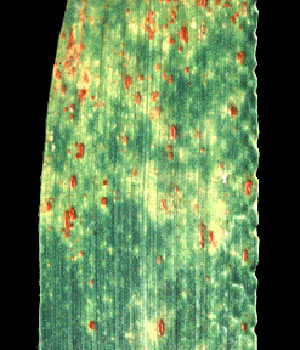
اختلافات بین گونه ها می تواند به جای اختلاف در متابولیسم, به ظرفیتهای متفاوت برای جذب و انتقال آب در یک وضعیت آبی مشخص منتهی گردد. تغییرات در نسبت ریشه به اندام هوایی یا تجمع موقتی ذخایر در ساقه تحت شرایط کمبود آب با تغییرات در متابولیسم کربن و نیتروژن همراهی می شود. در سطح برگ پراکندگی انرژی القایی (تهییجی) بوسیلهٔ فرایندهایی غیر از متابولیسم کربن فتوسنتزی یک مکانیسم دفاعی مهم می باشد که با کاهش در فتوشیمی و در دراز مدت افت ظرفیت فتوسنتزی و رشد توأ م می گردد.
● حفظ موازنهٔ صحیح آبی:
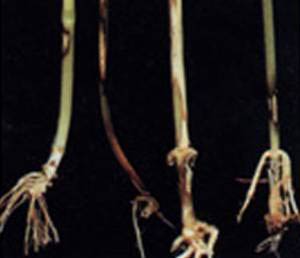
در همین رابطه, دو گونهٔ بلوط همیشه سبز را در کنار هم در منطقه اورای پرتقال, مورد مقایسه قرار داده و در یافتند که هیچ اختلاف معنی داری از نظر مقادیر آسمیلاسیون خالص کربن هنگامیکه رطوبت کافی در خاک وجود داشت و یا در مورد گیاهان تحت تنش خشکی ملایم در اول جولای, وجود ندارد. با وجود این, تا انتهای تابستان گرم و خشک (ماه سپتامبر) مبادلهٔ گازی نیمروزی در Q.ilex نسبت به Q. suberكمتر متأثر گردید. یعنی تا انتهای تابستان پتانسیلهای آبی بسیار بالاتری در برگهای Q.ilex (۵۲ .۱- مگا پاسگال ) در مقایسه با ۳۸ .۲-مگا پاسگال برای Q. suber مشاهده گردید. و چنین فرض شد که ریشه های Q.ilex قادر به مکش و دریافت آب از لایه های عمیق تر خاک بودند که به آنها اجازه می داد در مقایسه با Q. suber برای دورهٔ طولانی تری مقادیر بالاتری از جریان آب و آسمیلاسیون برگی را حفظ کنند. افزایش تراکم ریشه در واحد حجم خاک در گیاهان یک ساله نظیرLupinus albus نیز در شرایط کمبود آب مشاهده شده است در کل, رشد اندام هوایی در مقایسه با ریشه در برابر کمبود آب حساس تر است. مکانیسم هایی که زمینهٔ پایداری و تداوم رشد ریشه را تحت تنش خشکی فراهم می آورند شامل تنظیم اسمزی, افزایش در ظرفیت از بین رفتهٔ دیوارهٔ سلولی و همچنین انباشت ABA داخلی به جهت ممانعت از تولید اتیلن, می باشند.
▪ بسته شدن روزنه ,ذخیرهٔ اقتصادی آب برای آسمیلاسیون کربن:
كنترل روزنه ایی تلفات آب, که هم در واکنش به کاهش در تورژسانس برگی یا پتانسیل آبی و هم رطوبت نسبی پایین هوا می تواند اتفاق بیفتد, به عنوان اولین واکنش گیاه به کمبود آب در شرایط مزرعه ایی تشخیص داده شده است. بطوریکه روزنه ها به علایم شیمیایی (مثلABA) تولید شده بوسیلهٔ ریشه های دهیدراته پاسخ می دهند در حالیکه وضعیت آبی برگ ثابت نگه داشته می شود.
co۲قابل دسترس, کنترل کنندهٔ ظرفیت بیوشیمیایی آسمیلاسیون کربن می باشد که کاهش در کربن بین سلولی (Ci) به دنبال بسته شدن روزنه ها در دراز مدت کاهش ظرفیت ماشین فتوسنتزی را به منظور سازگاری به کربن در دسترس القا می کند.
بطوركلی, مقاومت به خشكی در گیاه به مجموعه ایی از مكانیسم ها و واكنش های پیچیده ایی گفته می شود كه گیاه در صورت بر خورد با كم آبی, توانایی رشد و نمو خود را تا حدودی حفظ می كند.
حسین خوشوقتی
منابع مورد استفاده:
۱- Chaves MM, Pereir JS, Maroco J, Rodriguse ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Pinheiro C. ۲۰۰۲. How plants cope with water stress in the field. photosynthesis and growth. annals of botany ۸۹: ۹۰۷-۹۱۶.
۲- Rodrigues ML, Pacheco CA, Chaves MM. ۱۹۹۵. Soil-plant relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. Journal of Experimental Botany ۴۶: ۹۴۷–۹۵۶.
۳- Sharp RE, LeNoble ME. ۲۰۰۲. ABA, ethylene and the control of shoot and root growth under water stress. Journal of Experimental Botany ۵۳: ۳۳–۳۷.
۴- Wilson KB, Baldocchi DD, Hanson PJ. ۲۰۰۱. Leaf age affects the seasonal pattern of photosynthetic capacity and net ecosystem exchange of carbon in a deciduous forest. Plant Cell and Environment ۲۴: ۵۷۱–۵۸۳.
منابع مورد استفاده:
۱- Chaves MM, Pereir JS, Maroco J, Rodriguse ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Pinheiro C. ۲۰۰۲. How plants cope with water stress in the field. photosynthesis and growth. annals of botany ۸۹: ۹۰۷-۹۱۶.
۲- Rodrigues ML, Pacheco CA, Chaves MM. ۱۹۹۵. Soil-plant relations, root distribution and biomass partitioning in Lupinus albus L. under drought conditions. Journal of Experimental Botany ۴۶: ۹۴۷–۹۵۶.
۳- Sharp RE, LeNoble ME. ۲۰۰۲. ABA, ethylene and the control of shoot and root growth under water stress. Journal of Experimental Botany ۵۳: ۳۳–۳۷.
۴- Wilson KB, Baldocchi DD, Hanson PJ. ۲۰۰۱. Leaf age affects the seasonal pattern of photosynthetic capacity and net ecosystem exchange of carbon in a deciduous forest. Plant Cell and Environment ۲۴: ۵۷۱–۵۸۳.




همچنین مشاهده کنید











































































































نمایندگی زیمنس ایران فروش PLC S71200/300/400/1500 | درایو …
دریافت خدمات پرستاری در منزل
pameranian.com
پیچ و مهره پارس سهند
خرید میز و صندلی اداری
خرید بلیط هواپیما
گیت کنترل تردد
اسرائیل ایران اصفهان ایران و اسرائیل حجاب فرانسه حمله ایران به اسرائیل انفجار حسین امیرعبداللهیان گشت ارشاد دولت ارتش جمهوری اسلامی ایران
سیل هواشناسی تهران فضای مجازی طرح نور وزارت بهداشت قتل شهرداری تهران زلزله سیلاب سازمان هواشناسی قوه قضاییه
قیمت طلا قیمت دلار دلار قیمت خودرو دولت سیزدهم خودرو بانک مرکزی بازار خودرو چین ایران خودرو حقوق بازنشستگان تورم
سعدی سینمای ایران سینما تلویزیون موسیقی فیلم احسان علیخانی دفاع مقدس کتاب تئاتر سریال
آزمون دکتری
رژیم صهیونیستی عراق غزه فلسطین جنگ غزه امیرعبداللهیان سازمان ملل شورای امنیت اسراییل عملیات وعده صادق حماس ترکیه
فوتبال استقلال پرسپولیس شمس آذر قزوین باشگاه استقلال لیگ برتر صنعت نفت آبادان لیگ قهرمانان اروپا بازی رئال مادرید تراکتور لیگ برتر فوتبال ایران
هوش مصنوعی فناوری گوگل سامسونگ ناسا تلگرام اپل وزیر ارتباطات عیسی زارع پور
حمله قلبی بارندگی فشار خون مغز درمان و آموزش پزشکی پزشک آلزایمر